I read this with a sense of wonderment:
New footprints from Laetoli (Tanzania) provide evidence for marked body size variation in early hominins (Open Access)
- Fidelis T Masao, Elgidius B Ichumbaki1, Marco Cherin, Angelo Barili,Giovanni Boschian, Dawid A Iurino, Sofia Menconero, Jacopo Moggi-Cecchi, Giorgio Manzi

Source: Fidelis T Masao et al 2016: Trackway L8 with four footprints (top). Relief map of the trackway L8 (bottom).
The tracks shown in the image above is the L8 trackway from site S, made by individual S1. Site S contains another set of tracks made by individual S2. Another site named site G, discovered some years earlier, lies about 150 meters to the north of site S. It contains tracks made by 3 individuals G1, G2 and G3. At both sites the tracks indicate that the hominins were walking in the same north- northeasterly direction. The tentative conclusion is that S1 is a large male, S2 and G2 are adult females and G1 and G3 are juveniles or smaller adult females. Based on fossils found in this stratigraphic succession it is believed that these tracks were made by individuals of the species Australopithecus afarensis.
Geological and stratigraphic reasoning lead to the conclusion that the trackways at site G and site S have been imprinted on the same tuff (volcanic ash) layer (Footprint Tuff) which contain a record of ash deposition over just a few weeks. Remarkably, this means that we could have a 3.66 million year old record of two contemporaneous trackways made by the same general population of hominins living in this area.
As the title of the paper suggests the important observation here is a marked body size variation interpreted from the morphology of the footprints. Below is a neat figure of stature estimates of hominins found from various localities in Africa ranging in age from 4 million years ago to 1 million years ago. The size range of A. afarensis overlaps that of later Homo.
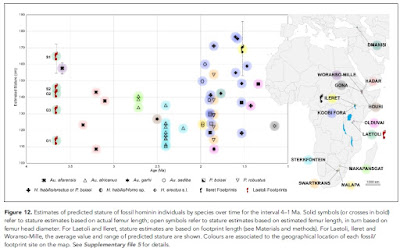
Source: Fidelis T Masao et al 2016
and an extract from the paper:
"These findings provide independent evidence for large body-size individuals among hominins as ancient as 3.66 Ma. Consequently, we may emphasise the conclusions by Grabowski et al. (2015) and Jungers et al. (2016), who reported that the body sizes of the australopithecines and of the early Homo representatives were similar, but also that certain australopithecine individuals (at least of Au. afarensis) were comparable with later Homo species, including H. erectus s. l. and H. sapiens. Thus, our results support a nonlinear evolutionary trend in hominin body size (Di Vincenzo et al., 2015; Jungers et al., 2016) and contrast with the idea that the emergence of the genus Homo and/or the first dispersal out of Africa was related to an abrupt increase in body size (McHenry and Coffing, 2000; Antón et al., 2014; Maslin et al., 2015). The identification of large-size individuals among the australopithecines – i.e. hominins commonly presumed to be small-bodied on average – shows also that the available fossil record can be misleading, resulting in an underestimate of the hominin phenotypic diversity in any given period".
.."Evidence for either marked or moderate body-size variation in Au. afarensis, based on data collected in a single site, was limited until now to the fossil assemblage from the Hadar 333 locality, dated to 3.2 Ma (with body masses ranging from 24.5 to 63.6 kg). The new estimates for the Laetoli individuals indicate an even more marked variation in body size within the same hominin population, at 3.66 Ma. Consequently, the combined records from Laetoli and Hadar suggest that large-bodied hominins existed in the African Pliocene for over 400,000 years, between 3.66 and 3.2 Ma. At the same time, these data contrast with the hypothesis of a temporal trend of body-size increase among Au. afarensis between the more ancient Laetoli and the more recent Hadar fossil samples (Lockwood et al., 2000)".
Another implication is that there was marked sexual dimorphism in Australopithecus afarensis possibly due to male to male competition. Their social structure may have been more similar to gorillas than chimpanzees or modern humans.
What a find!
New footprints from Laetoli (Tanzania) provide evidence for marked body size variation in early hominins (Open Access)
- Fidelis T Masao, Elgidius B Ichumbaki1, Marco Cherin, Angelo Barili,Giovanni Boschian, Dawid A Iurino, Sofia Menconero, Jacopo Moggi-Cecchi, Giorgio Manzi

Source: Fidelis T Masao et al 2016: Trackway L8 with four footprints (top). Relief map of the trackway L8 (bottom).
The tracks shown in the image above is the L8 trackway from site S, made by individual S1. Site S contains another set of tracks made by individual S2. Another site named site G, discovered some years earlier, lies about 150 meters to the north of site S. It contains tracks made by 3 individuals G1, G2 and G3. At both sites the tracks indicate that the hominins were walking in the same north- northeasterly direction. The tentative conclusion is that S1 is a large male, S2 and G2 are adult females and G1 and G3 are juveniles or smaller adult females. Based on fossils found in this stratigraphic succession it is believed that these tracks were made by individuals of the species Australopithecus afarensis.
Geological and stratigraphic reasoning lead to the conclusion that the trackways at site G and site S have been imprinted on the same tuff (volcanic ash) layer (Footprint Tuff) which contain a record of ash deposition over just a few weeks. Remarkably, this means that we could have a 3.66 million year old record of two contemporaneous trackways made by the same general population of hominins living in this area.
As the title of the paper suggests the important observation here is a marked body size variation interpreted from the morphology of the footprints. Below is a neat figure of stature estimates of hominins found from various localities in Africa ranging in age from 4 million years ago to 1 million years ago. The size range of A. afarensis overlaps that of later Homo.

Source: Fidelis T Masao et al 2016
and an extract from the paper:
"These findings provide independent evidence for large body-size individuals among hominins as ancient as 3.66 Ma. Consequently, we may emphasise the conclusions by Grabowski et al. (2015) and Jungers et al. (2016), who reported that the body sizes of the australopithecines and of the early Homo representatives were similar, but also that certain australopithecine individuals (at least of Au. afarensis) were comparable with later Homo species, including H. erectus s. l. and H. sapiens. Thus, our results support a nonlinear evolutionary trend in hominin body size (Di Vincenzo et al., 2015; Jungers et al., 2016) and contrast with the idea that the emergence of the genus Homo and/or the first dispersal out of Africa was related to an abrupt increase in body size (McHenry and Coffing, 2000; Antón et al., 2014; Maslin et al., 2015). The identification of large-size individuals among the australopithecines – i.e. hominins commonly presumed to be small-bodied on average – shows also that the available fossil record can be misleading, resulting in an underestimate of the hominin phenotypic diversity in any given period".
.."Evidence for either marked or moderate body-size variation in Au. afarensis, based on data collected in a single site, was limited until now to the fossil assemblage from the Hadar 333 locality, dated to 3.2 Ma (with body masses ranging from 24.5 to 63.6 kg). The new estimates for the Laetoli individuals indicate an even more marked variation in body size within the same hominin population, at 3.66 Ma. Consequently, the combined records from Laetoli and Hadar suggest that large-bodied hominins existed in the African Pliocene for over 400,000 years, between 3.66 and 3.2 Ma. At the same time, these data contrast with the hypothesis of a temporal trend of body-size increase among Au. afarensis between the more ancient Laetoli and the more recent Hadar fossil samples (Lockwood et al., 2000)".
Another implication is that there was marked sexual dimorphism in Australopithecus afarensis possibly due to male to male competition. Their social structure may have been more similar to gorillas than chimpanzees or modern humans.
What a find!